编辑 | 水产研究社
最新首发 | 了解最新水产养殖知识,关注:水产研究社
水产养殖,病害防治,水质调节,技术交流,生态养鱼,养虾,养蟹,健康渔业,您值得信赖的水产小帮手。

近年来,集约化的养殖模式推动了水产养殖产业的发展,但是在集约化的养殖模式下,鱼类易患营养性、代谢性及应激性疾病,这些问题也制约了水产养殖产业的发展,对水产养殖业造成了严重的经济损失。
乌鳢是典型的肉食性物种,因生长快、抗病力强和市场价值高等特点在我国被广泛养殖。

近年来,珠三角地区鳢科鱼类的养殖品种主要是杂交鳢,2021年珠三角地区杂交鳢的产量达到28.30万吨,占比全国总产量的51.60%。
淀粉是饲料配方中重要的膨化剂和黏合剂,在饲料制作过程中使膨化饲料达到物理膨胀。
然而,鱼类属于葡萄糖(GLU)不耐受体质,高碳水化合物饲料可诱发高血糖、糖原积累及炎症反应,进而损害肝脏免疫和代谢机能,最终导致鱼类肝脏代谢性疾病。
牛磺酸( taurine)首次从牛胆汁中分离出,因此被称为牛胆素。
牛磺酸由膳食摄取或自身合成,其中,水产品及畜禽产品中含量更为丰富,牛磺酸不参与蛋白质合成,在水产动物体内以游离氨基酸形式分布并发挥生理功能。
牛磺酸在水产动物体内主要用于维持细胞内外渗透压,清除氧自由基、提高机体免疫力、改善内分泌状态及促进糖、脂代谢等。
研究表明,饲料碳水化合物含量的提高可降低饲料中优质蛋白质的比例,由于优质蛋白质含量的下降,牛磺酸的含量也随之减少。
大黄鱼(Larimichthys cro.cea)和真鲷( Pagrus major)饲料缺乏牛磺酸会降低其生长性能,相反,牛磺酸的补充会改善这种现象,并提高饲料利用率。
另外,据报道牛磺酸可激活胰岛素信号通路,增强胰岛素敏感性并降低胰岛素抵抗等功能。
然而关于牛磺酸对杂交鳢饲喂高碳水化合物饲料的缓解作用和机制目前还鲜有报道。
因此,我们假设牛磺酸具有可缓解或抑制水生动物由高碳水化合物饲料引起的肝脏代谢性疾病。
长期摄入高碳水化合物饲料会抑制鱼类生长,导致肝糖原积累,进而诱导炎症反应和肝脏功能异常。
肉食性鱼类饲料碳水化合物的添加量通常为 20%左右,杂交鳢饲料碳水化合物含量超过 25%会导致高血糖和脂质氧化积累以及炎症反应,进而损害免疫和抗氧化能力。
饲料添加牛磺酸可以显著促进鱼类生长,提高免疫力,激活胰岛素信号通路,促进糖代谢。
日本牙鲆、鲈鱼、大菱鲆和尼罗罗非鱼等水产动物的研究表明,牛磺酸在饲料中的适宜添加量为1%左右。
然而,目前关于牛磺酸在杂交鳢饲料中应用的研究较为缺乏。
因此,本研究以杂交鳢为试验对象研究高碳水化合物饲料中添加牛磺酸对杂交鳢生长性能、糖代谢、肝脏抗氧化活性及免疫应答的影响及作用机制,旨在为牛磺酸在水产养殖中的应用提供依据。
1材料与方法
1.1试验饲料
试验设计3个组:低碳水化合物组(LC组,碳水化合物含量为 21%)、高碳水化合物组(HC 组碳水化合物含量为27%)及高碳水化合物饲料添加牛磺酸组(HT 组,碳水化合物含量为 27%,牛磺酸添加量为 1%)。
牛磺酸纯度≥98%。按照饲料配方将不同的饲料原料进行称量,并粉碎过 60 目筛,逐级混合后拌水,采用B20强力搅拌机(广州市番禺力丰食品机械厂)进行混合,混合后采用T52 型膨化机(广州市华强膨化机械厂)制成膨化饲料,继而将磷脂油、豆油和鱼油进行混合并喷洒到膨化饲料上,喷洒均匀后自然风干,风干后阴凉保存备用。试验饲料组成及营养水平见表1。

1.2试验鱼和养殖管理
在广东省农业科学院动物科学研究所白云试验基地进行养殖试验,养殖用水需曝气后使用。从广州市锦龙渔业有限公司购买杂交鳢鱼苗。

第1周在 2.5 mx2.5 mx1.5 m的暂养网箱中暂养,每天 08:00 和 16:00 饱食投喂LC 组饲料2次。
暂养结束后禁食24h,随机挑选体格均匀、初始体重为(22.02±0.02)g、活力旺盛的杂交鳢鱼苗450尾,随机分为3组,每组3个重复,每个重复50尾,以重复为单位在 1.5 mx1.5 mx1.5 m 的网箱中进行8周养殖试验。
每天08:00 和16:00 饱食投喂各组试验饲料2次。试验期间养殖水体溶氧含量在 8 mg 左右,pH 8.0左右,氨氮含量<0.1 mg/L,水温为 25~32 ℃,试验采取自然光照。
1.3 数据分析
试验数据用平均值+标准误表示,采用SPSS25.0软件进行统计分析,先对试验数据进行方差齐性检验,满足方差齐性条件则进行单因素方差分析(one-way ANOVA),差异显著再用 Tukey’s检验方法进行多重比较:方差齐性条件不满足,则用 Dunnett’s T3 检验法进行多重比较。差异显著性水平为 P<0.05。
2结果
2.1生长性能分析
由表3可知,与HC组相比,HT组FC显著降低(P<0.05).PPV 显著升高(P<0.05);各组间杂交鳢FBW、WGR、SR和SGR无显著差异(P>0.05)。

2.2血浆生化指标分析
由表4 可知,与 HC 组相比,HT 组血浆 TP 含量显著升高(P<0.05);
与LC 组和 HT 组相比,HO组血浆 GLU 含量显著升高(P<0.05);
与 LC 组相比,HC 组和HT 组血浆 AST 活性显著升高(P<0.05):;
与LC 组相比,HC 组和 HT 组血浆 ALP 活性显著下降(P<0.05);
与LC 组和 HT 组相比,HC组血浆 ALT活性显著升高(P<0.05);
各组间血浆TG、UN 和 TCHO 含量无显著差异( P>0.05)。
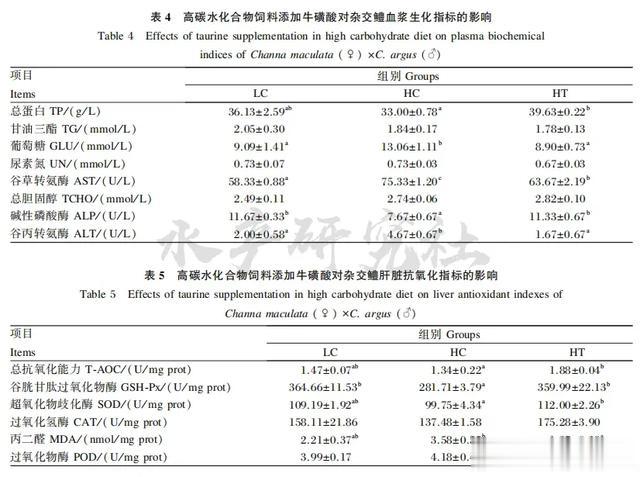
2.3 肝脏抗氧化指标分析
由表5 可知,与 HC 组相比,HT 组肝脏T-AOC显著升高(P<0.05);
与 HC 组相比,LC 组和 HT组肝脏 GSH-Px 活性显著升高(P<0.05);
与HC组相比,HT 组肝脏 SOD 活性显著升高(P<0.05);
与HC组相比,HT组肝脏MDA含量显著升高(P<0.05);
各组肝脏 CAT 和 POD 活性无显著差异(P>0.05)。
2.4肝脏糖原和胰岛素含量分析
由表6 可知,与 HC 组相比,HT 组肝脏的肝糖原含量显著下降(P<0.05);
与LC组和HC组相比,HT 组肝脏胰岛素含量显著升高(P<0.05);各组间肝脏肌糖原含量无显著差异(P>0.05)。

2.5肝脏胰岛素信号通路相关基因表达分析
由图1 可知,与 HC 组相比,HT 组肝脏蛋白激酶B(AKT)和磷脂酰肌醇-3-羟激酶( PI3K)基因相对表达量显著上调(P<0.05);
各组肝脏胰岛素受体底物-1(IRS-1)基因相对表达量无显著差异(P>0.05)。
2.6肝脏糖代谢相关基因表达分析
由图 2 可知,与 LC 组和 HC 组相比,HT 组肝脏丙酮酸激酶(PK)、葡萄糖激酶(GK)和葡萄糖转运体2(GLUT2)基因相对表达量显著上调(P<0.05);

各组间肝脏糖原合成酶(GS)磷酸烯醇丙酮酸羧激酶( PEPCK)和葡萄糖-磷酸酶(G6P)基因相对表达量无显著差异(P>0.05)。
2.7肝脏免疫相关基因表达分析
由图 3 可知,与 HC 组相比,HT 组肝脏白细胞介素-8(IL-8)基因相对表达量显著下调(P<0.05);
与 HC 组相比,LC 组和 HT 组肝脏白细胞介素-1B(IL-1B)和肿瘤坏死因子-α(TNF-a)基因相对表达量显著下调(P<0.05);
与 HC 组相比、LC 组和HT 组肝脏转化生长因子-B(TGF-8)基因相对表达量显著上调(P<0.05);
与 HC 组相比,HT组肝脏热休克蛋白70(HSP70)基因相对表达量显著上调(P<0.05);
与LC 组和 HC 组相比,HT组肝脏热休克蛋白90(HSP90)基因相对表达量显著上调(P<0.05);
与 HC 组相比,HT 组肝脏白细胞介素-10(IL-10)基因相对表达量显著上调(P<0.05);
与 HC 组相比,LC 组和 HT 组肝脏 Toll 样受体4(ILR4)和髓样分化因子88(MyD88)基因相对表达量显著下调(P<0.05)。

2.8肝脏病理切片分析
由图 4 可知,LC 组和 HT 组杂交鳢肝索结构清晰,肝细胞排列紧密,脂质含量丰富,表现为胞浆空白,部分静脉周围可见肝胰脏,细胞形态正常,组织未见明显坏死或炎症反应。
HC组杂交肝胰脏周围间隙增宽(黑色箭头)小叶偶见炎性细胞小灶性浸润(红色箭头),未见其他明显异常。

讨 论
肉食性鱼类对饲料碳水化合物利用率较低通常控制在 20%以下。
当饲料碳水化合物含量过高会导致鱼类生长抑制、肝糖代谢异常及炎症反应。
Xu 等研究表明,杂交鳢碳水化合物含量为11%能够显著促进生长、提高抗氧化活性和免疫应答。
本研究表明,高碳水化合物饲料降低了杂交鳢 WGR、SGR 和 PPV,升高FC,但并不显著,这说明高碳水化合物饲料对杂交鳢产生生长抑制作用。
本研究结果与大黄鱼研究结果相似。饲料碳水化合物含量较高情况下鱼类会通过高采食量维持正常的生长性能,这就解释了杂交鳢采食高碳水化合物饲料导致的FC高但生长性能降低的原因。
在高碳水化合物饲料中添加牛磺酸后,杂交鳢 FC 显著下降,PPV 显著提高,WGR 和 SGR也相应提高,但并不显著,说明添加牛磺酸可显著提高杂交鳢的生长性能。
同时牛磺酸能够提高PPV,减少蛋白质作为能源物质损耗。
牛磺酸还可刺激鱼类采食感受器,增强摄食效率,这充分解释了杂交鳢FC显著下降,生长性能显著提高的原因,同时也在牙鲆和大菱鲆等多种鱼种中得到验证。
血浆生化指标关联到鱼类的营养代谢及健康状况。
在本试验条件下,杂交鳢摄食高碳水化合物饲料后,血浆 GLU含量显著升高。
这可能由于在鱼类体内进行营养代谢过程中,高碳水化合物饲料会导致鱼类出现持续高血糖现象。这与虹鳟和欧洲海鲈研究结果相似。
当高碳水化合物饲料添加牛磺酸后,牛磺酸表现出良好的降血糖效果,这可能由于牛磺酸可以促进肌细胞摄取和利用 GLU,加速糖酵解,降低血糖浓度,也可以诱导糖原合成酶I活性升高,增加糖原的合成,同时降低糖原磷酸化酶活性,抑制糖原分解。
同时观察到,饲料添加牛磺酸后,血浆TP含量显著升高,这与杂交鳢 PPV结果一致,表明饲料添加牛磺酸可以显著促进机体蛋白质生成,这可能由于牛磺酸可以促进大量的含硫氨基酸合成蛋白质。
因此,可以推测出牛磺酸可以促进杂交的蛋白质合成及代谢。
血清代谢包括ALTAST和AKP,是检测动物健康状况的重要血清标志物,ALT AST、AKP等组织标记解活性的改变与器官特异性细胞损伤程度有直接关系。
ALP被认为参与生长蛋白质合成,营养物质的摄取和运输等。
在本试验中观察到高碳水化合物饲料饲喂杂交鳢导致血浆 AST 和ALT 活性显著升高转氨酶活性升高代表了鱼类健康受到高碳水化合物饲料的影响。
在高碳水化合物饲料中添加牛磺酸后,杂交鳢血浆 ALT 活性显著下降,ALP 活性显著升高,这说明添加牛磺酸可保护肝脏功能免受高碳水化合物饲料损害,显著改善杂交鳢的健康状况。
机体氧化反应产生自由基,进而损坏机体细胞,引起疾病的发生。
机体细胞内存在氧化防御体系,可以清除氧化反应产物,使机体保持氧化动态平衡状态。
机体氧化防御体系包括酶系统(SOD、GSH-Px、T-AOC)和非解系统(维生素E和维生素 C))。
T-AOC 反映机体抗氧化能力SOD反映机体清除氧自由基的能力.GSH-P反映机体清除过氧化氢的能力,MDA反映组织间脂质氧化损伤的程度。
在本试验条件下,高碳水化合物饲料显著降低了杂交鳢肝脏GSH-P的活性.SOD活性和T-AOC也出现降低,但不显著。
本试验结果表明,高碳水化合物饲料可降低机体的抗氧化能力,这可能由于高碳水化合物饲料降低了肝脏 SOD 活性,机体氧自由基显著增加,肝脏发生氧化损伤,T-AOC显著下降。
同时观察到,当在高碳水化合物饲料中添加牛磺酸后,杂交鳢肝脏 T-AOC、GSH-Px 和 SOD 活性显著升高,MDA 含量显著下降,对杂交鳢肝脏抗氧化能力的改善尤其明显,这表明高碳水化合物饲料中添加牛磺酸可以缓解杂交鳢的氧化应激,增强机体抗氧化能力,这可能由于牛磺酸可与自由基结合,阳止氧化反应的进行,直接发挥抗氧化作用。
此外,牛磺酸还可调节线粒体蛋白质合成,促进线粒体呼吸活动,进而保护线粒体,增强机体抗氧化活性,这与石首鱼和欧洲鲈鱼研究结果相似。
持续性高碳水化合物饲料的摄入,会使机体出现高血糖现象,这种现象也被认为机体胰岛素分泌不足。
在本试验条件下,HC组的杂交肝脏胰岛素含量低于LC组,并表现出高血糖现象。
本试验结果表明,杂交鳢吸收到体内的糖类不能很好的利用,因此杂交鳢对糖类的利用效率可能与胰岛素的分泌效率相关。
同时观察到,在高碳水化合物饲料中添加牛磺酸可显著提高肝脏胰岛素含量,这可能由于牛磺酸能抑制胰腺 ATP敏感的钾离子通道活性,使 KATP 通道关闭,β 细胞去极化激活电压依赖性的钙离子(Ca2+)通道增加细胞内Ca2+,从而刺激胰岛素分泌,进而促进细胞摄取和利用 GLU,加快糖酵解效率,达到降血糖作用。
IRS-1是胰岛素通路(IRS-1/PI3K/AKT)的第一信使,当IRS-1激活后,胰岛素信号会进入细胞并激活 P3K,进而通过 AKT行使胰岛素样作用。
在本试验条件下,HC组杂交鳢肝脏AKT和PI3K的基因表达水平被显著抑制,HT组肝脏AKT和PI3K基因相对表达量显著上调,这也有力地证明了牛磺酸对杂交鳢表现出的胰岛素样作用,牛磺酸进入胰腺β细胞能够引起胰岛素的释放,从而使血浆 GLU 含量降低,以调控杂交鳢的糖代谢过程。
GLU 代谢的第1步是GLUT2 介导 GLU 和肝细胞之间的相互转运。
PK和GK在糖酵解中发挥限速作用,在本试验中,高碳水化合物饲料抑制了PK和GLUT2 的表达,这种抑制会直接影响到GLU在肝脏和血液之间的转运速率,这一点从肝脏糖原累积可以表明高碳水化合物饲料抑制了糖原分解效率,从而造成肝糖原蓄积,影响肝脏糖代谢。
牛磺酸的添加显著促进PK和GK的基因表达,说明牛磺酸可以促进糖酵解并提高肝脏糖代谢效率。
肝脏对免疫系统的贡献非常重要,炎症反应作为鱼类免疫系统反应的重要组成部分,在应对各种挑战方面做出了巨大贡献。
炎症反应是先天免疫系统应对各种刺激的重要组成部分,主要是由细胞因子介导(IL-8、IL-1β、TNF-α、TGF-β和IL-10)。
IL-8主要促进中性粒细胞黏附在内皮细胞上并迁移到炎症部位,IL-1β和TNF-α可以通过调节其他细胞因子的表达来诱导炎症反应,TGF-β通过抑制炎性细胞因子的释放来有效缓解炎症,IL-10 抑制T淋巴细胞的增殖 、活化和迁移,限制炎症反应。
HSP 在促进身体细胞对各种压力的抵抗力方面起着重要作用,因此常用于水生动物转录调控和应激反应的研究。
HSP70和HSP90基因表达水平的上调可以显著增强机体细胞对各种应激的抵抗力,保护细胞免受氧化应激。
研究表明,摄入高碳水化合物饲料可引起肝脏损伤,脂肪沉积和炎症反应等。
在本试验条件下,高碳水化合物饲料显著上调了肝脏IL-1β和INF-α基因相对表达量,显著下调了TGF-β和HSP70基因相对表达量。
本试验结果表明,肉食性鱼类由于肝糖应激,饲料中高碳水化合物直接损伤肝脏,造成肝细胞损伤及炎症反应。
同时观察到,在高碳水化合物饲料中添加牛磺酸肝脏 IL-8 IL-1β,TNF-α、TLR4 和MyD88 基因相对表达量显著下调,1L-10、TGF-β和HSP70基因相对表达量显著上调。
这说明高碳水化合物饲料中添加牛磺酸可以显著改善杂交鳢的免疫能力。
TLRS 是一类跨膜蛋白,具有识别入侵病原体和触发细胞信号转导等功能,MyD88 和TLR4 参与类症过程。
在本研究中,高碳水化合物饲料添加牛磺酸显著下调了杂交鳢肝脏中MYDB8和TLR4基因的表达,结果表明,牛磺酸可激活杂交鳢的Tol 样受体,介导MyD88依赖的信号通路,最终抑制促炎细胞因子IL-8、IL-1β 和 TNF-α 的释放激活获得性免疫反应,最终调节免疫功能,增强机体抵抗力,减轻肝脏炎症反应。
由此可见,饲料中添加牛磺酸可以通过调节抗氧化酶的活性来保护肝脏免受氧化损伤,通过上调抗类因子的表达和抑制促炎因子的表达来减轻肝脏炎症反应。
在本试验条件下,当杂交鳢在高碳水化合物应激状态肝脏进入病理状态,进而表现出HSP70基因相对表达显著下调,随着牛磺酸的添加,肝脏MyDE和TLR4 基因相对表达量显著上调,TLRS 通过MyD88依赖通路诱导免疫应答。
综合分析表明牛磺酸可通过调节杂交鳢肝脏 TLR4-MyD88 信号通路相关因子减轻炎症反应,改善免疫功能。
肝脏组织形态被认为是评价鱼类营养状况代谢和肝脏健康的良好指标。
为了评估补充牛磺酸对高碳水化合物饲料引起的组织病理学改的有效性,制作了肝脏切片。
先前的研究表明,高碳水化合物饲料会损害肝脏,例如组织间隙增宽炎症反应及细胞核破裂等。
在本研究的高碳水化合物组中也可以观察到炎性细胞浸润症状。同样,在本研究中,饲料中添加牛磺酸减少了由高碳水化合物饲料引起的肝脏组织学异常。
由此可见,饲料中添加牛磺酸可缓解杂交鳢高碳水化合物饲料所诱导的肝脏炎症病理状态。
4 结 论
综上所述,高碳水化合物饲料的摄入诱导了氧化应激、炎症反应、糖代谢素乱甚至肝脏形态改变,显著降低了杂交鳢的生长性能。
高碳水化合物饲料中添加牛磺酸显著降低了FC,这可使杂交鳢养殖饲料成本降低。
此外,在高碳水化合物饲料中添加牛磺酸提高了杂交鳢肝脏的抗氧化能力,促进了肝脏胰岛素信号通路(PI3K和AKT)的激活,并通过调控 TLR4-MyD88 免疫信号通路,抑制促炎症因子(IL-1β 和TNF-α)基因表达,缓解肝脏炎症反应。
