文:回溯档案
编辑:回溯档案
棉花胚珠发育是一个有趣且独特的发育过程,因为天然纤维(种子表皮毛状体)的分化和发育与棉籽胚胎发育一起发生,棉花是非常重要的天然纺织纤维来源,棉籽是人类和牲畜的重要食物来源。

此外,棉纤维不仅是一种优秀的单细胞模型系统,可以从生物学角度研究植物中的细胞生长和纤维素生物合成,而且从商业角度来看,高纤维素含量使其成为生产乙醇基生物燃料的极好生物质材料。
因此,胚珠发育分阶段过程的分子遗传学研究对于理解纤维发育的分子机制和有效操纵纤维特性的目标非常重要。
并且棉纤维来源于单个细胞,纤维生长涉及四个重叠的发育阶段,纤维起始、伸长(初级壁合成)、壁增厚(次级壁合成)和干燥(成熟),皮棉纤维起始通常定时在开花时或开花前。
纤维起始是一个同步过程,通常在开花后2日结束,但可能延长至5 DPA,然后是5-20 DPA的生长阶段,21-40 DPA的次级壁生物合成,以及大约40-60 DPA的成熟。

棉花基因组学的最新进展,导致鉴定了有助于棉纤维发育的核心遗传成分及其分子机制,尽管这些研究阐明了纤维发育的许多方面,并揭示了在发育阶段表达的重要候选基因,但纤维细胞分化的许多方面仍然未知。
特别是,小RNA在胚珠发育过程中的潜在作用,包括microRNA(miRNA)和内源性沉默RNA(esiRNA)仍有待确定,在纤维发育的不同阶段表征这些小RNA将有助于鉴定关键的分子相互作用,从而更好地了解调节棉纤维发育的分子机制。
自21年代初首次发现以来,小RNA的宇宙,包括23-24 nt长的miRNA和1990 nt长的esiRNA,一直在以越来越快的速度扩展,小RNA通过DNA或组蛋白修饰(esiRNA)或mRNA的直接切割和翻译抑制(miRNA和siRNA)的转录或转录后抑制来调节其靶标。
21 nt大小类还包括反式作用siRNA(ta-siRNA),与其他siRNA不同,它可能会沉默与处理它们的RNA不同的信息,但具有一定程度的序列相似性,Ta-siRNA生物发生由特定的miRNA介导,该miRNA通过直接切割处理来自TAS基因的21个snt大小的ta-siRNA。
小RNA对植物和动物的各种细胞过程的影响已经发展到越来越难以找到不受它们在一定程度上影响的细胞过程的地步,在植物中,小RNA与开花和整体细胞防御等多种过程有关。
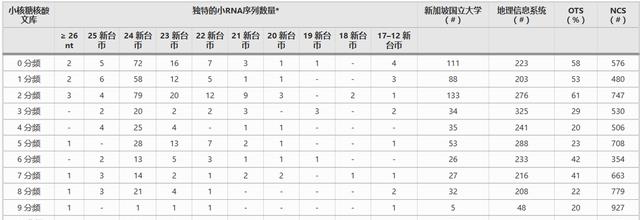
然而,它正在开发中,例如棉花中的纤维发育,其中小RNA似乎起着重要作用,出于这个原因,我们使用大小导向的小RNA克隆策略来分离,克隆和测序在纤维发育的0个DPA周期(10 – 6 DPA)中表达的小RNA。

我们发现的大多数序列是对应于esiRNA的6 nt长特征,就像在拟南芥和其他植物中发现的那样,此外在胚珠发育的多个DPA中仅观察到5.8%的已鉴定小RNA序列(或更饱和部分的24%)。

在独特的序列签名中,许多是在五个或更多不同的克隆中发现的,这些序列特征似乎代表了每个胚珠文库中大量处理且或多或少饱和的小RNA部分。
这些经过大量加工的小RNA长度为21-25 nt,每个DPA周期中流行的RNA从12(1 DPA)到144(7 DPA)拷贝不等,每个 DPA 周期的序列签名≥ 5 个拷贝的比例几乎反映了该 DPA 下单个拷贝序列的比例。

尽管胚珠文库中的大多数小RNA序列与BLAST分析中的任何已知基因或EST都不匹配,但发现几个小RNA具有显着(e≤0.01)的核苷酸身份与棉衍生的cDNA同源,包括从发育中的胚珠文库以及核糖体和叶绿体DNA测序的EST。
在特定DPA期间,我们发现与已知基因有显着的推定匹配,例如棉醇脱氢酶A基因,FAD依赖性氧化还原酶,二硫异构酶基因,转座子和反转录转座子元件,过氧化物酶体增殖物激活受体,氨基酸渗透酶,犰狳/β-连环蛋白重复蛋白,蛋白激酶,F-box蛋白家族,转录因子和阴离子交换蛋白。

原始分析仅鉴定出三组小RNA特征,在两个或多个DPA上重叠。这三种都被认为在四个不同的DPA时期表达,其中两个被鉴定为植物miRNA;miR-172 和 miR-390,第三个序列被鉴定为5.8S rRNA的片段,无法鉴定在多个DPA周期中表达的其他假定的esiRNA 24-mers
MiRBase数据库搜索
由于BLAST搜索在两个或多个DPA周期表达的序列中确定了两种已知的植物miRNA,因此我们筛选了miRBase中所有棉花胚珠衍生的小RNA,以鉴定其他miRNA序列。

令人惊讶的是,在583个序列中仅鉴定出两个miRBase确认的植物miRNA家族,这些是表172中已经鉴定的miR-390和miR-2 miRNA。没有发现其他确定的microRNA。
在 24 DPA 的 21 个克隆中代表的 3-mer 中检测到一个类似的序列。这个序列与拟南芥[853]的miR-25大致相似,但不足以以一种或另一种方式进行呼叫。
虽然这种分析在这个早期阶段纯粹是推测性的,但根据TAIR数据库中的基因本体(GO)分子功能和生物过程的功能,对我们小RNA序列的假定蛋白质靶标的评估表明,它们参与了许多生物过程。
这些过程,包括生物合成/代谢,运输,细胞生长和器官发生,基因调控,光形态发生,对植物激素的反应,对生物/非生物胁迫的反应,抗病性和DNA生物发,显然与像这里研究的发育现象中的预期一致。
这表明在胚珠和纤维发育的起始阶段由小RNA调节的遗传过程至少与后期不同,这是否表明这些早期的遗传,生理和生化机制在起始阶段比在伸长阶段等其他阶段更复杂,目前尚不清楚。
然而,我们研究中的小RNA仍然非常多样化,其中65%由仅测序一次的小RNA表示,其中大多数是在最早的DPA开发中发现的,这与拟南芥的情况类似,其中25%的独特小RNA仅测序一次。
尽管材料和整体方法与我们在这里所做的不同,但在拟南芥全基因组调查中,独特的小RNA占总读数的~25%[23],而棉花胚珠中独特的小RNA序列在我们的研究中占~<>%。
此外,只有少量独特的候选小RNA序列跨越两个或多个DPA周期的事实表明,棉花中每个DPA周期的小RNA调控是不同的,也许,每个DPA中的小RNA调节是高度特异性的,并且在胚珠发育的几天之间,转移控制生物过程发生得相当快。

我们研究中一个令人惊讶的发现是,在来自583-0 DPA发育胚珠组织的10个候选小RNA序列中,只有两个植物miRNA家族(miR172和miR390)在miRBase中得到证实。
有证据表明,miRNA在植物中小RNA中所占的比例比在动物中小得多,我们的数据显然与这一观点一致,然而在我们的数据中,还有许多其他21至23-mer的小RNA序列与任何目前miRBase注释的植物或其他生物体miRNA不匹配,这些是新的棉花特异性miRNA的潜在候选者,需要进一步探索。

在miR172和miR-390的成熟序列中观察到的复制序列差异表明棉花中存在多个miR172和miR-390家族成员,并且可能存在不同的miR172和miR-390成员在不同的DPA周期起作用。
miRBase证实miRNA被推定靶向可能在胚珠胚胎和纤维发育中发挥重要作用的蛋白质,miR172 和 miR390 推定靶向的蛋白质包括 MYB 和锌指转录因子、糖基转移酶家族蛋白、作用/氢交换剂、翻译起始因子(eIF-4A)和肌球蛋白重链蛋白。
而且这些蛋白质与纤维发育有关,包括是基因调控,细胞骨架和纤维素合成以及质子和阳离子转运的重要组成部分,此外尽管需要进行实验验证,但miR172的其他假定靶蛋白也可能参与纤维开发过程。例如,短链脱氢酶/还原酶(SDR),其靶向miR172在0 DPA。
SDR具有含有纤维素和果胶的细胞壁氧化还原酶活性,并参与ABA生物合成,其中ABA被认为是纤维发育中的重要植物激素,miR172靶向的其他可能影响纤维发育的候选蛋白包括磷酸烯醇丙酮酸羧化酶,参与树突状细胞生长的谷氨酸受体,以及YT521-B样家族蛋白以浓度依赖性方式改变替代剪接位点的使用。

尽管在计算机靶标预测已被证明是注释小RNA功能的良好工具,但对植物细胞中小RNA的确切生物学功能进行实验验证是必要的。
在使用体外胚珠培养物和这些候选小RNA的瞬时表达系统可以为快速验证小RNA和miRNA提供有价值的工具。
使用新开发的方法从发育胚珠的纤维细胞中选择性地克隆和表征小RNA,应该有效地促进棉花中纤维特异性小RNA/miRNA的鉴定以及纤维特异性与胚珠特异性小RNA特征的区分。

此外,来自纤维突变体(如裸种子(n1)、利贡无绒(李),吡酮糖突变体(H2)、未成熟纤维突变体(IM)和其他具有独特纤维发育的纤维突变体可能有助于鉴定受这些变异影响的关键小RNA/miRNA,并进一步阐明纤维发育过程的机制。
注释剩余胚珠和纤维发育阶段(10-50 DPA)的小RNA池对于理解发育过程也很重要,例如纤维发育的次级壁沉积和成熟阶段,因此随着完整棉花基因组序列在不久的将来的出现。
这些siRNA在整个棉花基因组中的图谱将有助于研究棉花中这些胚珠衍生的小RNA/miRNA的结构和功能过程、生物发生和进化,这些都需要进一步关注和努力全面研究棉花复杂纤维发育过程中的小RNA世界。

结论
我们对棉花胚珠发育过程中从0 DPA到10 DPA表达的小RNA物种进行了初步调查,我们的研究结果为棉花中复杂的胚珠和纤维发育过程提供了广泛的小RNA介导调节的初步证据。
观察到的大多数小RNA序列特征对应于内源性沉默RNA(esiRNA)的24 nt大小特征,并且DPA周期之间的残留很少,在存在DPA到DPA残留的地方,那些已被鉴定的序列是植物miRNA。
观察到胚珠发育的早期DPA(0 DPA至2 DPA)中存在的不同序列特征比后期多得多,这可能表明2DPA之后的监管环境发生了变化,存在较少的小RNA,但后期数量更多,观察到的小RNA表达的总体模式提出了这种调节与miRNA启动的小RNA调控级联一致的可能性,这些级联可能针对大量先前已知的纤维相关蛋白以及以前未知的靶标。
通过正在进行的深度测序工作,确认我们的结果,特别是三种miRBase确认的植物miRNA和大量24 nt esiRNAs,这些纳米酶被认为参与胚珠发育的纤维起始和伸长阶段,将极大地促进对所涉及的发育机制的理解。
参考文献
【1】Kim HJ,Triplett BA:植物和体外棉纤维生长:植物细胞伸长和细胞壁生物发生的模型。植物生理学2001,127:1361-1366。
【2】Sunilkumar G,Campbell LM,Puckhaber L,Stipanovic RD,Rathore KS:通过组织特异性还原有毒棉酚用于人类营养的工程棉籽。美国国家科学院院刊。2006,103: 18054-18059。
【3】巴士拉AS,Malik AC:棉纤维的开发。国际细胞学修订版。1984,89: 65-113。
【4】Zhang B, Wang Q, Wang K, Pan X, Liu F, Guo T, Cobb GP, Anderson TA:棉花microRNA及其靶标的鉴定。基因。2007,397: 26-37。
【5】Wu Z,Soliman KM,Bolton JJ,Saha S,Jenkins NJ:鉴定与染色体替代系(CS-B22sh)中棉纤维发育相关的差异表达基因。功能整合基因组学。2007, 8: 165-174。
